David R. Liu
David R. Liu
碱基编辑技术的出现,使得科幻小说中的构想有望成为令人激动的现实,未来我们能够给予孩子的最重要的礼物,可能不仅仅是30亿个DNA字母,还有保护和修复它们的方法。
人类单倍体基因组含有约31.6亿个DNA碱基对,碱基对是以氢键相结合的两个含氮碱基,以胸腺嘧啶(T)、腺嘌呤(A)、胞嘧啶(C)和鸟嘌呤(G)四种碱基排列成碱基序列,其中A与T之间有两个氢键配对,G与C之间有三个氢键配对。碱基的点突变是引发人类遗传病的主要原因之一,据ClinVar database显示,由C•G到T•A的突变,约占已知人类致病单核苷酸突变的一半。如果可以在基因组DNA中将目标A•T碱基对转化为G•C碱基对,将有可能纠正大部分与人类疾病相关的SNP。[1, 4]
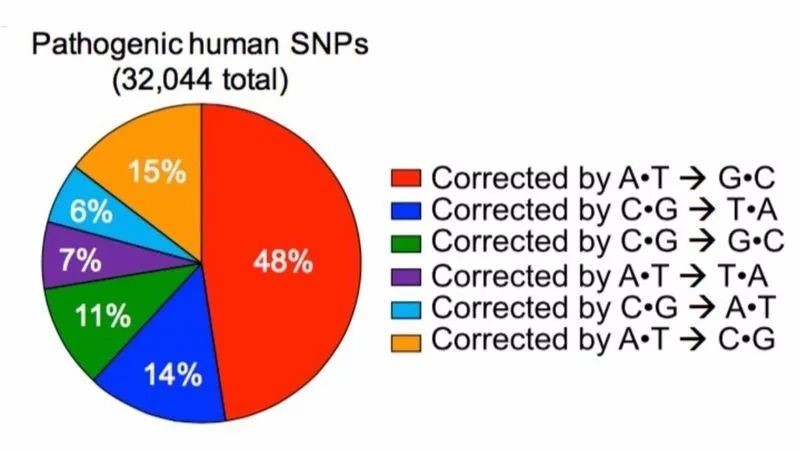
图1:引发人类遗传病的单碱基突变
近年来CRISPR技术(Clustered regularly interspaced short palindromic repeats)被广泛应用于各种生物的基因组编辑。然而,如果将CRISPR技术用于基因治疗或生物定向改造,需要对突变位点的精准识别,不能出现意外的随机插入或者缺失。美国哈佛大学David R. Liu课题组发表在《Nature》的” Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage”论文创新性的开发了基于CRISPR/Cas9的单碱基编辑器(base editor, BE),有望为上述问题提供新的思路。
相比传统的CRISPR/Cas9系统,单碱基基因编辑技术具有以下优势[2]:
单碱基编辑器可以将单核苷酸突变(SNV)引入活细胞的DNA或者RNA中,是基因组编辑领域的最新进展之一。 单碱基编辑器可以分为两大类:针对DNA的和针对RNA的。科学家们不断的改造和优化单碱基编辑器,使得DNA和RNA单碱基编辑器都取得了重要发展。
DNA碱基编辑器分为嘧啶碱基编辑器(cytosine base editors, CBEs)和嘌呤碱基编辑器(adenine base editors, ABEs)两种类型,分别适用于C/G到T/A和A/T到G/C的碱基对转变。CBEs 和 ABEs工作原理如图2所示,DNA碱基编辑器是向活细胞中引入永久性DNA点突变的有效工具。
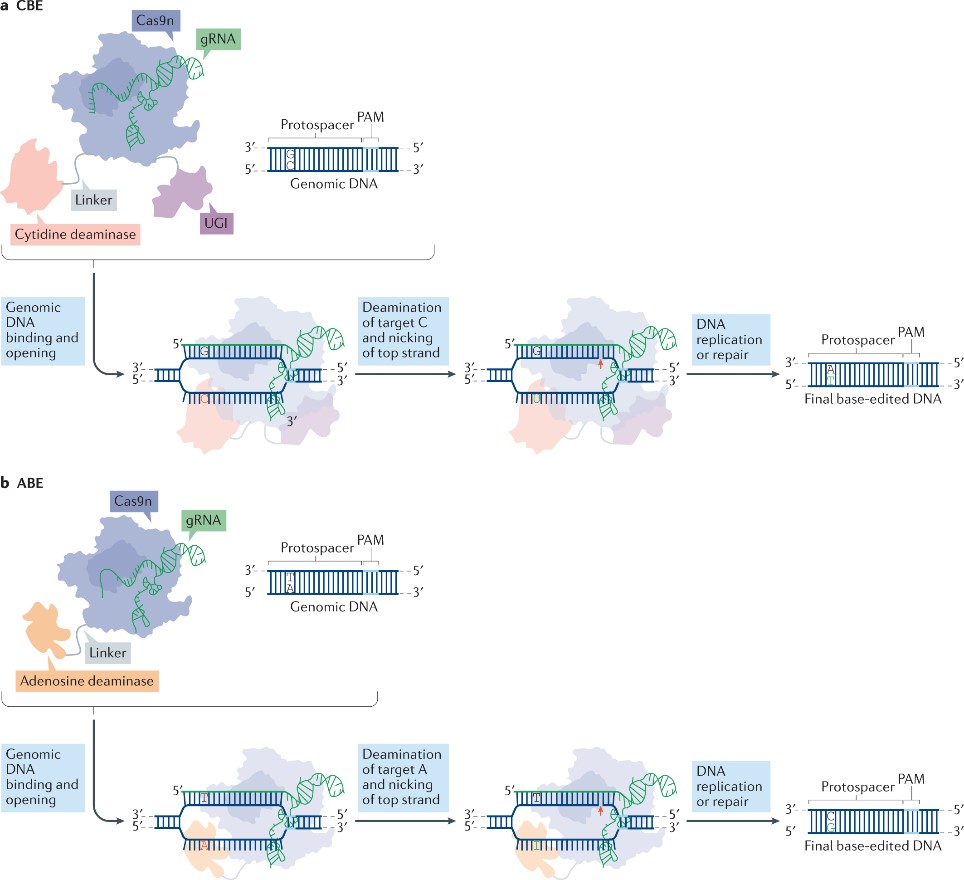
图2. DNA碱基编辑技术概述[3-4] a. 嘧啶碱基编辑器机制;b. 嘌呤碱基编辑器机制。
RNA碱基编辑器分为A到I碱基编辑器和C到U碱基编辑器,并根据其引入的修饰进一步细分。与DNA碱基编辑器不同,这些修饰在转录本的生命期内不会被细胞进一步处理。RNA碱基编辑器工作原理如图3所示。
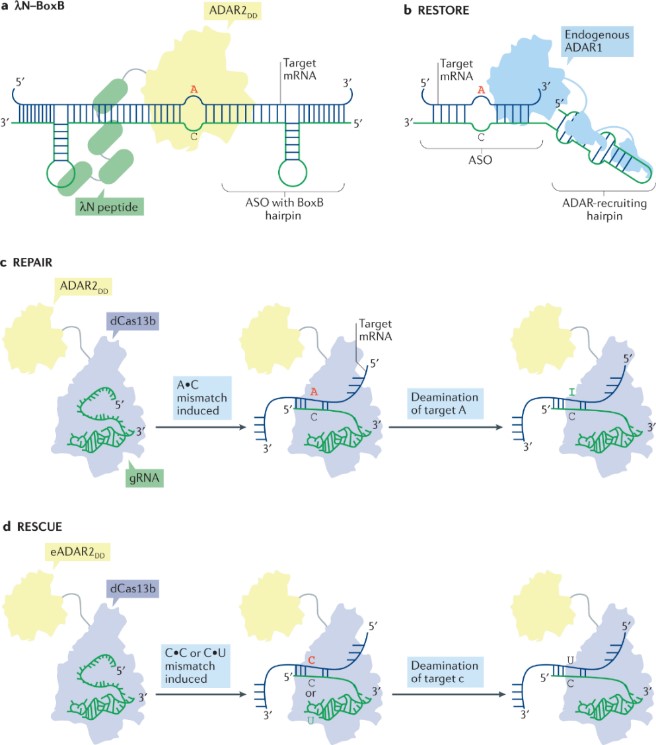
[5-8] a-c. A到I RNA碱基编辑;d. C到U RNA碱基编辑
2020年10月19日,加州大学欧文分校的研究人员在 Nature子刊 Nature Biomedical Engineering 杂志发表了题为:Restoration of visual function in adult mice with an inherited retinal disease via adenine base editing 的研究论文[9]。该研究使用慢病毒载体递送腺嘌呤单碱基编辑器(ABE),实现了对先天性黑蒙症小鼠模型突变基因的高效修复,有效恢复了小鼠模型视觉能力,且未发现可检测的脱靶效应。这项工作表明,单碱基编辑技术在某些情况下可能会替代基因增强疗法,以永久挽救因突变而失能的关键蛋白的功能,或修正无法使用基因增强疗法的显性遗传病。这项工作也代表了治疗遗传性视网膜疾病的新方向。
2020年10月19日,加州大学欧文分校的研究人员在 Nature子刊 Nature Biomedical Engineering 杂志发表了题为:Restoration of visual function in adult mice with an inherited retinal disease via adenine base editing 的研究论文[9]。该研究使用慢病毒载体递送腺嘌呤单碱基编辑器(ABE),实现了对先天性黑蒙症小鼠模型突变基因的高效修复,有效恢复了小鼠模型视觉能力,且未发现可检测的脱靶效应。这项工作表明,单碱基编辑技术在某些情况下可能会替代基因增强疗法,以永久挽救因突变而失能的关键蛋白的功能,或修正无法使用基因增强疗法的显性遗传病。这项工作也代表了治疗遗传性视网膜疾病的新方向。
韩国首尔大学Jin-Soo Kim组率先发表了将 rAPOBEC-XTEN-Cas9n-UGI( BE3) 系统运用于小鼠疾病模型制备的研究成果[11]。针对小鼠抗肌萎缩蛋白基因Dmd设计sgRNA,通过显微注射小鼠受精卵,得到9只F0代小鼠中,有5只小鼠产生了靶位点的碱基突变,包括一只纯合子突变小鼠( CAG>TAG) ;针对小鼠酪氨酸酶基因Tyr设计sgRNA,通过电转染方法,得到7只F0后代小鼠中均检测到靶基因突变,包括3只纯合子Tyr突变小鼠。中山大学黄军就组也利用单碱基编 辑系统研究了该技术在制备小鼠疾病模型中的效率[12]。并首次报道了在小鼠胚胎中由碱基编辑系统诱导产生的靶位点处的碱基插入以及临近位点的脱氨基。
中科院的高彩霞团队率先将单碱基编辑技术应用于三大重要农作物小麦、水稻和玉米的性状改良上,并且取得了重大进展[13]。高教授团队构建了密码子优化的 rAPOBEC1-XTEN-dCas9n-UGI 和 rAPOBEC1-XTEN-dCas9-UGI 两种融合蛋白,通过针 对三大农作物共 7 个基因位点进行单碱基编辑实验,发现选用 Cas9n 可以获得较高的单碱基编辑效率。同期中科院上海生命科学研究院的朱健康团队和中国农业科学院作物研究所的夏兰琴团队也分别报道了利用单碱基编辑系统对水稻基因进行单碱基编辑[14, 15]。单碱基编辑系统可以在农作物中取得高效的编辑效率,为改良作物品种带来新希望。[16]
本研究中使用的引物池由金斯瑞合成。引物池合成服务一次性合成多达91,766条引物,序列覆盖率99%以上,碱基单价低至1分钱,让您不再为预算和周期而烦恼,专心进行合成生物学研究、靶向测序及复合DNA文库实验等,将您的所有科研设想变成现实。